LUZ VISIBLE

Lo que comúnmente llamamos luz es una onda electromagnética y aunque tal vez, esto sea algo difícil de imaginar, como cualquier onda tiene una “longitud de onda”, es decir, la distancia que mide un ciclo completo de la onda, de forma que a partir de ahí, vuelve a repetirse lo mismo una y otra vez: Cada longitud de onda corresponde a lo que nosotros identificamos como un color. Ahora bien, nosotros no somos capaces de ver la luz de cualquier longitud de onda, nuestros ojos están limitados a ver desde 380 a los casi 780 nanometros ( 1 nanometro es una milésima de millonesima de metro). A esta franja se la llama “espectro visible”: Si a esta franja la llamamos espectro visible, cualquiera podría percatarse de que existe la insinuación de que hay un espectro no visible, algo así como “luz invisible”. Esto es cierto, pero hablaré de ello en un próximo artículo.
EL OJO Y LAS CÉLULAS RECEPTORAS
Principales segmentos de un cono o bastón:
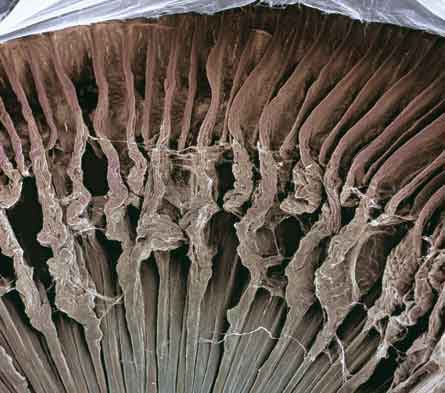 1. Segmento externo: aquí es donde se encuentra el fotopigmento sensible a la luz. En el caso de los bastones la rodopsina y en el caso de los conos los pigmentos del color.
1. Segmento externo: aquí es donde se encuentra el fotopigmento sensible a la luz. En el caso de los bastones la rodopsina y en el caso de los conos los pigmentos del color.
2. Segmento interno:
3. Núcleo.
4. Cuerpo sináptico. Los conos presentan una estructura cónica, con sus núcleos alineados en una sola capa justo por debajo de la membrana limitante externa.
Sus segmentos internos y externos se proyectan dentro del espacio subretinal hacia el epitelio pigmentario. A nivel de la fóvea, donde sólo existen conos, sus cuerpos celulares se sitúan oblicuamente con respecto a sus procesos. Los bastones por otra parte poseen una morfología alargada con sus segmentos internos y externos rellenando el espacio entre los conos y los procesos de las células del epitelio pigmentario. Los cuerpos celulares de los bastones constituyen el resto de la capa nuclear externa, donde se sitúan formando varias capas. Aunque no siempre resulta claro en las preparaciones histológicas habituales, los procesos de las células del epitelio pigmentario rodean completamente tanto a los procesos externos de los conos como de los bastones. Utilizando el microscopio electrónico se obtiene una mejor resolución de la morfología de los conos y bastones. Podemos observar cómo los segmentos internos de los bastones son más delgados que los de los conos. Así los segmentos internos de los conos poseen un diámetro de unas 6 micras frente a las 2 micras de los segmentos internos de los bastones. Sin embargo a nivel de la fóvea, donde sólo existen conos, los segmentos internos de éstos pueden llegar a medir tan sólo alrededor de 1.5 micras. Los segmentos internos de los conos y bastones poseen muchas mitocondrias. A nivel de la unión entre los segmentos internos y externos de los fotorreceptores existe un cilio de unión. A partir de estos cilios se producen una serie de evaginaciones e invaginaciones de la membrana plasmática de los fotorreceptores que dan lugar a los segmentos externos (o.s. en la imagen). Esta es la porción de los fotorreceptores donde se encuentran los pigmentos visuales. Los segmentos externos de los conos y bastones derivan de repliegues de la membrana plasmática (ver las figuras que se adjuntan a continuación).
Al final a nivel de los bastones, los segmentos externos están constituidos por discos membranosos aislados de la membrana plasmática donde se encuentran inmersos los pigmentos sensibles a las radiaciones luminosas. Por contra en los conos no existen discos membranosos aislados sino múltiples repliegues de la membrana plasmática.
 En los bastones, la molécula de GMPc actúa como segundo mensajero y transporta la información a través del citoplasma situado entre los discos que flotan libremente en la zona donde se absorbe la luz, hasta la membrana plasmática de la célula donde se produce la alteración de los flujos iónicos. En los conos debido a la continuidad entre los discos y la membrana plasmática no es necesario un mensajero citoplasmático. En estas células la molécula de GMPc controla los flujos de iones, abriendo unas clases especiales de canales iónicos, los canales iónicos regulados por el GMPc, permitiendo que una corriente de entrada que transporta sobre todo iones sodio, penetre en la célula. En la oscuridad, la concentración de GMPc es relativamente elevada, lo que determina que los canales iónicos regulados por esta molécula se encuentren abiertos y permitan la corriente de entrada que mantiene a la c‚lula en un estado relativamente despolarizado. Por tanto, la fotoconversión tiene lugar en tres fases: La luz activa los pigmentos visuales. Estos pigmentos visuales activados estimulan la GMPc fosfodiesterasa, una enzima que reduce la concentración de GMPc en el citoplasma. La reducción de la concentración de GMPc cierra los canales regulados por él, hiperpolarizando al fotorreceptor. Pasemos ahora al estudio detallado de cada una de estas fases. Veamos cómo los pigmentos visuales de las células fotorreceptoras transforman una señal luminosa en un impulso nervioso. Este proceso se ha estudiado fundamentalmente en los bastones, responsables de la visión escotópica (nocturna), que no es en color. Por su parte los conos se encargan de la visión fotópica (diurna), que se produce con color.
En los bastones, la molécula de GMPc actúa como segundo mensajero y transporta la información a través del citoplasma situado entre los discos que flotan libremente en la zona donde se absorbe la luz, hasta la membrana plasmática de la célula donde se produce la alteración de los flujos iónicos. En los conos debido a la continuidad entre los discos y la membrana plasmática no es necesario un mensajero citoplasmático. En estas células la molécula de GMPc controla los flujos de iones, abriendo unas clases especiales de canales iónicos, los canales iónicos regulados por el GMPc, permitiendo que una corriente de entrada que transporta sobre todo iones sodio, penetre en la célula. En la oscuridad, la concentración de GMPc es relativamente elevada, lo que determina que los canales iónicos regulados por esta molécula se encuentren abiertos y permitan la corriente de entrada que mantiene a la c‚lula en un estado relativamente despolarizado. Por tanto, la fotoconversión tiene lugar en tres fases: La luz activa los pigmentos visuales. Estos pigmentos visuales activados estimulan la GMPc fosfodiesterasa, una enzima que reduce la concentración de GMPc en el citoplasma. La reducción de la concentración de GMPc cierra los canales regulados por él, hiperpolarizando al fotorreceptor. Pasemos ahora al estudio detallado de cada una de estas fases. Veamos cómo los pigmentos visuales de las células fotorreceptoras transforman una señal luminosa en un impulso nervioso. Este proceso se ha estudiado fundamentalmente en los bastones, responsables de la visión escotópica (nocturna), que no es en color. Por su parte los conos se encargan de la visión fotópica (diurna), que se produce con color.

Cada color primario es percibido por una zona determinada de la retina. Hay cuatro zonas dispuestas en forma de anillos concéntricos con suaves límites de transición. De adentro hacia fuera son: la zona del verde, la del rojo, la del azul y la del blanco. La zona del verde percibe todos los colores por lo que decimos que es tricromática; la segunda es dicromática porque no percibe el verde y la última solamente capta a este. Otra particularidad del ojo es su aberración cromática: la luz blanca, al refractarse, desvía en forma desigual a los rayos que la componen; los que tienen menos longitud de onda son los que más se inclinan y forman su foco mas adelante al atravesar una lente. Los rayos mas cortos establecen su foco por delante y los más largos por detrás. La aberración cromática se corrige en parte por la actividad del iris que estrecha la abertura pupilar y elimina los rayos del margen del cristalino, que son los que más se desvían.
1. Segmento externo: aquí es donde se encuentra el fotopigmento sensible a la luz. En el caso de los bastones la rodopsina y en el caso de los conos los pigmentos del color.2. Segmento interno:
3. Núcleo.
4. Cuerpo sináptico. Los conos presentan una estructura cónica, con sus núcleos alineados en una sola capa justo por debajo de la membrana limitante externa.
Sus segmentos internos y externos se proyectan dentro del espacio subretinal hacia el epitelio pigmentario. A nivel de la fóvea, donde sólo existen conos, sus cuerpos celulares se sitúan oblicuamente con respecto a sus procesos. Los bastones por otra parte poseen una morfología alargada con sus segmentos internos y externos rellenando el espacio entre los conos y los procesos de las células del epitelio pigmentario. Los cuerpos celulares de los bastones constituyen el resto de la capa nuclear externa, donde se sitúan formando varias capas. Aunque no siempre resulta claro en las preparaciones histológicas habituales, los procesos de las células del epitelio pigmentario rodean completamente tanto a los procesos externos de los conos como de los bastones. Utilizando el microscopio electrónico se obtiene una mejor resolución de la morfología de los conos y bastones. Podemos observar cómo los segmentos internos de los bastones son más delgados que los de los conos. Así los segmentos internos de los conos poseen un diámetro de unas 6 micras frente a las 2 micras de los segmentos internos de los bastones. Sin embargo a nivel de la fóvea, donde sólo existen conos, los segmentos internos de éstos pueden llegar a medir tan sólo alrededor de 1.5 micras. Los segmentos internos de los conos y bastones poseen muchas mitocondrias. A nivel de la unión entre los segmentos internos y externos de los fotorreceptores existe un cilio de unión. A partir de estos cilios se producen una serie de evaginaciones e invaginaciones de la membrana plasmática de los fotorreceptores que dan lugar a los segmentos externos (o.s. en la imagen). Esta es la porción de los fotorreceptores donde se encuentran los pigmentos visuales. Los segmentos externos de los conos y bastones derivan de repliegues de la membrana plasmática (ver las figuras que se adjuntan a continuación).
Al final a nivel de los bastones, los segmentos externos están constituidos por discos membranosos aislados de la membrana plasmática donde se encuentran inmersos los pigmentos sensibles a las radiaciones luminosas. Por contra en los conos no existen discos membranosos aislados sino múltiples repliegues de la membrana plasmática.
FOTOQUÍMICA DE LA VISIÓN
La retina es la parte del ojo sensible a la luz, que contiene los conos y los bastones, fotorreceptores responsables de la visión en color y en la oscuridad respectivamente. Nuestro objetivo en este estudio es comprender la transformación de una señal luminosa en impulso nervioso gracias a la actuación de los pigmentos visuales de las células fotorreceptoras (conos y bastones). La absorción de luz por los pigmentos visuales de los bastones ( rodopsina) y los conos activa una cascada de fenómenos bioquímicos que tiene lugar a través de la membrana plasmática de estas células y, por tanto, un cambio del potencial de membrana. Una molécula esencial en esta cascada es el nucleótido cíclico 3’- 5’ monofosfato de guanosina (GMPc).En los bastones, la molécula de GMPc actúa como segundo mensajero y transporta la información a través del citoplasma situado entre los discos que flotan libremente en la zona donde se absorbe la luz, hasta la membrana plasmática de la célula donde se produce la alteración de los flujos iónicos. En los conos debido a la continuidad entre los discos y la membrana plasmática no es necesario un mensajero citoplasmático. En estas células la molécula de GMPc controla los flujos de iones, abriendo unas clases especiales de canales iónicos, los canales iónicos regulados por el GMPc, permitiendo que una corriente de entrada que transporta sobre todo iones sodio, penetre en la célula. En la oscuridad, la concentración de GMPc es relativamente elevada, lo que determina que los canales iónicos regulados por esta molécula se encuentren abiertos y permitan la corriente de entrada que mantiene a la c‚lula en un estado relativamente despolarizado. Por tanto, la fotoconversión tiene lugar en tres fases: La luz activa los pigmentos visuales. Estos pigmentos visuales activados estimulan la GMPc fosfodiesterasa, una enzima que reduce la concentración de GMPc en el citoplasma. La reducción de la concentración de GMPc cierra los canales regulados por él, hiperpolarizando al fotorreceptor. Pasemos ahora al estudio detallado de cada una de estas fases. Veamos cómo los pigmentos visuales de las células fotorreceptoras transforman una señal luminosa en un impulso nervioso. Este proceso se ha estudiado fundamentalmente en los bastones, responsables de la visión escotópica (nocturna), que no es en color. Por su parte los conos se encargan de la visión fotópica (diurna), que se produce con color.VISION A COLOR
La visión de los colores es función del ojo adaptado a la luz y depende de los conos retinianos. Cuando la iluminación se reduce los objetos pierden el color según un orden determinado: rojo, amarillo, verde y azul. En oscuridad los colores dejan de percibirse y solo se distinguen matices, de gris, negro y blanco azulado; esta información es suministrada por los bastoncillos.Cada color primario es percibido por una zona determinada de la retina. Hay cuatro zonas dispuestas en forma de anillos concéntricos con suaves límites de transición. De adentro hacia fuera son: la zona del verde, la del rojo, la del azul y la del blanco. La zona del verde percibe todos los colores por lo que decimos que es tricromática; la segunda es dicromática porque no percibe el verde y la última solamente capta a este. Otra particularidad del ojo es su aberración cromática: la luz blanca, al refractarse, desvía en forma desigual a los rayos que la componen; los que tienen menos longitud de onda son los que más se inclinan y forman su foco mas adelante al atravesar una lente. Los rayos mas cortos establecen su foco por delante y los más largos por detrás. La aberración cromática se corrige en parte por la actividad del iris que estrecha la abertura pupilar y elimina los rayos del margen del cristalino, que son los que más se desvían.
PERCEPCIÓN
Puedes entar a la siguiente página: http://www.educacionplastica.net/ilusiones.htm, y aprenderás las teorias propuestas por personas preparadas y cocer tus capacidades visuales.
No hay comentarios:
Publicar un comentario